


Utilizing nationwide motion databases to assist inform responses to swine illness outbreaks in Scotland: the influence of uncertainty round incursion time
Impact of uncertainty in infection time
We looked at the extent to which inaccuracy in defining the disease incursion date may impact on the accuracy of predictions of pre-detection spread of acute swine diseases via movements of pigs. A premises-based model was developed to simulate their spread through the Scottish swine industry via movements of pigs, in which gathering places (such as markets and collection centres) were explicitly modelled together with pig producers. In the first instance, we considered the extreme case where infection occurs if at least 1 animal from an infected premises is received by a susceptible one. In this situation, the “infection paths” Γt,i of farms that were infected via movements of animals from a single pig producer i was computed for each Monday of the year 2012. Here, we considered all ith producers that were active during the period [t0, t0+T] eligible to be an index-case, where t0 is the incursion date and T is the “pre-detection period” (that is, the period between the date of the incursion t0 and the date of the first detection of the disease). We then compared the infection paths Γt,i with those Γt+δ,i generated when time of infection t0 is inaccurately estimated by an error δ ranging from −7δ0 to 7δ0. In this study, infection path Γt,i refers to the “correct” full epidemic tree that is generated by a single infection event at time t0 and left freely spreading for the pre-detection period [t0, t0+T], while Γt+δ,i refers to the “predicted” full epidemic tree when the incursion date is inaccurately estimated and for which the pre-detection period is [t0+δ, t0+T–δ]. We considered, δ0 = 7 days and T = 60 days19.
In Fig. 1, we show how increasing uncertainty around the time of incursion may affect one’s ability to accurately predict not only the number of premises involved in the full epidemic tree but also their identity. Overall, progressively increasing the error δ around the time of the incursion from δ0 to 7δ0 yielded a marked reduction in the correlation between sizes (i.e., the number of premises involved in) of infection paths Γt,i and Γt+δ,i (Fig. 1a,b). Although this reduction was consistent across paths of all sizes (Fig. 1a), it was more pronounced for paths of larger sizes (Fig. 1b). Also, there was a clear divide between infection paths generated from commercial producers and those generated from non-commercial producers (Fig. 1c). Despite a wide uncertainty on the time of the incursion, the correlation remained high between infection paths generated by commercial producers (Spearman’s correlation coefficient ρ > 0.60), whether assured or non-assured, for errors ranging from −7δ0 to 4δ0. In contrast, correlation between infection paths becomes weaker for incursions in non-commercial producers, with ρ < 0.60 for errors of ±3δ0.
Figure 1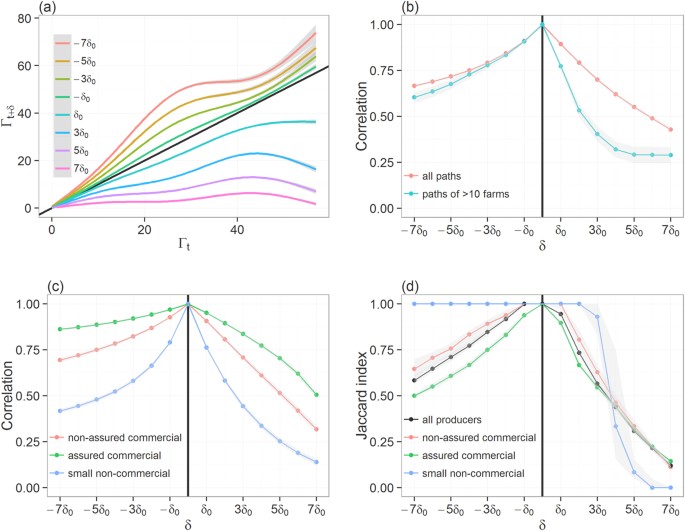
Comparison between the correct infection path and predicted paths generated when the error δ in the time of the incursion ranges from −7δ0 to 7δ0.
(a) Lines plot showing the smoothed size of the predicted full epidemic tree Γt+δ,i as a function of the size of the correct full epidemic tree Γt,i. (b) Changes in the Spearman correlation coefficient between the size of Γt,i and that of Γt+δ,i as a function of the error δ in the time of the incursion. Correlation coefficients are computed either upon all generated infection paths or upon infection paths of >10 infected premises. (c) Changes in the Spearman correlation coefficient between the size of Γt,i and that of Γt+δ,i as a function of δ and stratified by the producer type of the index-case. (d) Quality of infection path prediction, as measured by the median Jaccard similarity index, as a function of δ and stratified by the producer type of the index-case. Shaded areas around each line shown in (a–d) represent their respective confidence interval. Here, δ0 = 7days. Diagonal solid line in (a) indicates perfect concordance between the true and predicted length of infection paths. The vertical solid line in (b–d) indicates the time of the correct incursion time.
In order to see if we could accurately predict which individual premises would be involved in epidemics despite some inaccuracy in the incursion time, we compared the concordance between infection paths Γt,i and Γt+δ,i generated from the same index-case i, by calculating the Jaccard similarity index J(Γt,i,Γt+δ,i). The Jaccard index measures the fraction of common premises within paths |ξt,i∪ ξt+δ,i| among the total number of premises |ξt,i ∩ ξt+δ,i| involved in both paths. Here, we only focused on infection paths involving more than 10 infected premises.
Progressively increasing the error around the infection time up to 7δ0 revealed a reduction in the median degree of overlap between paths (Fig. 1d). The rate of this reduction differed, however, whether the incursion time is believed earlier (i.e. δ < 0) or later (i.e. δ > 0) than the true one. Overall, an error of −4δ0 in the infection time yielded 77% (95% CI 0.76–0.79) overlaps between the true and predicted paths, whereas an error of >2δ0 is enough to create completely different paths with paths involving, on average, less than half of common premises.
Unsurprisingly, variations between producer types were observed in the degree of overlap between Γt,i and Γt+δ,i. While the degree of overlap between predicted and the true paths generated by commercial producers followed closely the general trend, it differs greatly when considering paths generated by non-commercial small producers. This was expected, because most paths of >10 infected premises have been generated by commercial producers. However, differences between the degree of overlap for paths generated by commercial and those by non-commercial producers depends on the direction of the error δ: when δ < 0, predicted paths generated by non-commercial producers have a greater number of common premises with the true path, whereas paths would show a completely different pattern (i.e. J(Γt,i,Γt+δ,i) < 10%) from >4δ0 (Fig. 1d). These results suggest that if incursion occurs in non-commercial producers, conservative estimates in incursion times would be preferential. However, this may not be true for incursion occurring in commercial producers as a trade-off may exist between optimising the proportion of premises that are truly on the infection path (true positives) and minimising the proportion of premises that are not (false positives). Figure 2 explores how these two epidemiological measures vary with δ for paths generated by the different producer types. Over-estimating incursion times for outbreaks generated from commercial producers (whether assured or not), would increase the risk of misclassification. For example, inferences generated for outbreaks from non-assured and from assured commercial producers when δ = −5δ0 would involve 24% (95% CI 22–29%) and 39% (95% CI 37–41%) of false positives, respectively (Fig. 2b).
Figure 2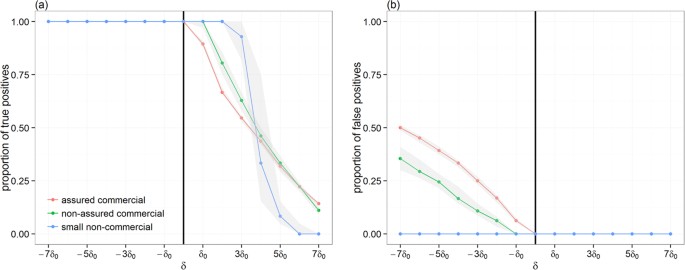
Proportions of true (a) and false (b) positives between the correct full infection path and predicted paths generated when the error δ in the time of the incursion ranges from −7δ0 to 7δ0.
Here, δ0 = 7days. Points/lines represent the observed median proportions, stratified as a function of the producer type of the index-case, whereas shaded areas represent their respective 95% confidence interval. Only infection paths of >10 infected premises are used. The vertical solid line indicates the time of the correct incursion time. The proportion of true positives measures the fraction of common premises within paths |ξt,i∪ ξt+δ,i| among the number of premises |ξt,i| that are on the correct path. The proportion of false positives measures the fraction of uncommon premises within paths 1 −|ξt,i∪ ξt+δ,i| among the number of premises |ξt+δ,i| that are on the wrong path.
So far in this analysis, the potential for spread of infection via movements of animals has been evaluated considering that any movement from infected premises during the pre-detection period would result in disease transmission to susceptible farms. In reality, the prevalence of disease within infected premises will determine what proportion β of its livestock becomes infectious. This, together with the number of animals that are being moved off, will determine what proportion of movements will contain infectious animals. To gain general insight and ensure robustness of the results to variation in β, 10,000 simulations for each Monday of the year 2012 with a random index-case per simulation were carried (i.e., total of 570,000 simulations). For each incursion date t0, the infection paths Γ’t,i of farms that were infected via animal movements from a single pig producer i was then computed and compared to the infection paths Γ’t+δ,i that were predicted when an error δ around the time of the incursion is made. As above, Γ’t,i and Γ’t+δ,i are the “correct” and “predicted” partial epidemic tree, respectively and correspond to all farms that have a non-null probability of being infected via animal movements from a single pig producer i. Figure 3 shows that, whether comparison is made with the “correct” full epidemic tree Γt,i (i.e. when β = 1) or with the “correct” partial epidemic tree Γ’t,i (i.e. when β < 1), qualitatively similar results as in Fig. 1 are obtained. However, it further appears that decreasing the value of β would reduce the effect of δ when predicting the size of the infection path (Fig. 3a). It is to note, however, that this result may give a false sense of security as the degree of overlaps between correct and predicted paths still sharply decreases with increasing error δ around the time of the incursion from δ0 to 7δ0 (Fig. 3b).
Figure 3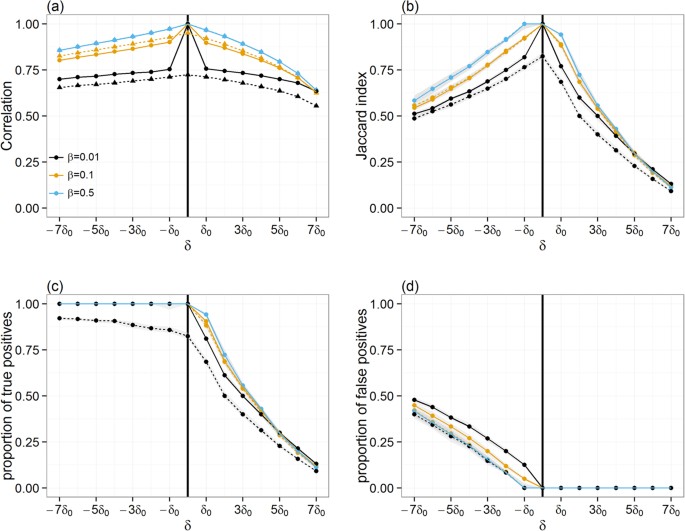
Comparison between the correct infection path and paths generated when the error δ in the time of the incursion ranges from −7δ0 to 7δ0 and when imperfect transmission occurs.
(a) Changes in the Spearman correlation coefficient between the size of the correct infection path and paths generated when both the error δ in the time of the incursion and the transmission probability β vary. Quality of infection path prediction, as measured by the median Jaccard similarity index (b), proportion of true (c) positives and proportion of false positives (d) between the correct infection path and paths generated when both δ and β vary. Solid and dotted lines indicate how measures may change when comparing predicted partial infection path Γ’t+δ,i to either the correct partial epidemic tree Γ’t,i generated with δ = 0 and β < 1 (solid) or the correct full epidemic tree Γt,i generated with δ = 0 and β = 1 (dotted). Shaded areas around each line shown in (b–d) represent the confidence interval around the median. Here, δ0 = 7days. The vertical solid line in (a–d) indicates the time of the correct incursion time.
Intrinsic structure of infection paths
Although our findings suggest that inferring the spread of an epidemic from dynamic network models is precarious when the date of the disease incursion is unknown, infection paths may have some intrinsic structure which may still guide contact tracing procedures. Previously, such a structure was found in the Italian cattle industry by comparing epidemic trees and regrouping index-cases which generated similar trees, thereby providing critical information to optimize surveillance systems and define rapid containment strategies4. Applying a similar method for the Scottish swine industry, however, would only result in regrouping producers that belong to the same business or are part of the same breeding pyramid. Instead, we looked at the producer type of both the index case and all farms that have been infected via the movement of animals when considering β = 1 and determined, for all full epidemic tree Γt,i that gave rise to at least 10 cases from the year 2012, the proportion of producers of each type that were involved in each infection path. The results are summarized in Fig. 4.
Figure 4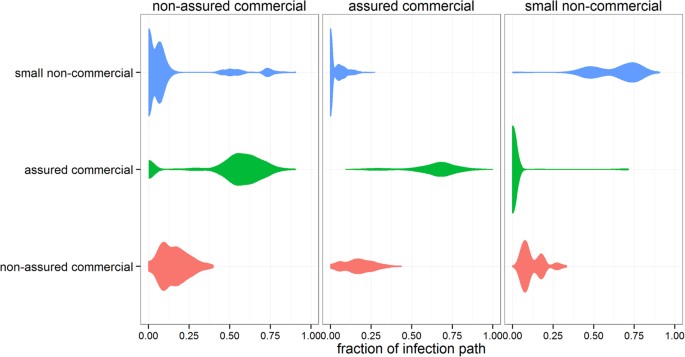
Proportion of non-assured commercial, assured commercial and small non-commercial producers involved in infection paths of >10 infected premises generated by each producer type.
Here, columns indicate the producer type of the index-case, whereas rows indicate the type of the producers that are involved in each infection path. The thickness of the shapes is proportional to the density of data points along the x-axis.
If disease incursion occurs in the herd of a small producer, the mean risk of disease spillover into assured producers is low (0.011); and similar to the mean risk of disease spillover from assured producers to small producers (0.032). Epidemics which start in a small producer spread into at least one assured producer in only 1.9% of the incursions. However, once an assured producer is infected, 60% (Q1–Q3: 17–71%) of the premises in the generated infection paths would belong to assured producers. In contrast, epidemics generated from assured producers would spread into small producers in 39% of the incursions, but would not involve many of them, with only 8% (Q1–Q3: 2–27%) of premises in these infection paths belonging to small producers. These findings are the consequence of producers adhering to quality assurance scheme guidelines on risks associated with animal trading20, confirming that excluding interactions with producers that have lower biosecurity standards is a good biosecurity practice21. Such a result may constitute a basis for the development of qualitative rules modulating surveillance activities in the face of an epidemic.
Non-assured commercial producers appear to have a totally different epidemiological profile (Fig. 4). Non-assured commercial producers have a consistently high probability (>95%) of being on an infection path and make up, on average, 17% (Q1–Q3: 9–22%) of premise in these paths, regardless of the producer type of the incursion. In addition, epidemics generated by non-assured producers show a high likelihood of infecting both small producers (0.62) and assured producers (0.85). This result highlights that Scottish swine producers who are commercially driven but do not belong to assurance schemes may potentially represent “epidemiological” bridges between non-commercial and commercial partners, likely because they implement lower biosecurity, particularly with regard to sourcing and sending pigs, compared to assured commercial producers. Therefore, improving biosecurity and targeting surveillance to non-assured producers may be particularly beneficial to optimise responses to disease incursions.
Comments are closed.